研究内容の概要
1.高等植物細胞のオルガネラ
高等植物は、細胞内に存在するオルガネラ(細胞内小器官)の機能を大きく変化させることで環境に適応しています。たとえば、脂肪性種子の細胞にはプロテインボディやオイルボディ、グリオキシソームなどのオルガネラが存在しています。これらは貯蔵物質の蓄積や分解を担うオルガネラで、種子が吸水した後に従属栄養成長を行う上で重要な役割を果たします。発芽が進行する過程では、オルガネラの機能変換や消失、形成などが起き、やがて葉緑体やミトコンドリア、緑葉ペルオキシソーム、液胞などが発達することで細胞が光合成能を獲得します。このように種子発芽過程におけるオルガネラの変化は、植物が従属栄養成長から独立栄養成長へと転換する上で重要な役割を果たしています(図1)。
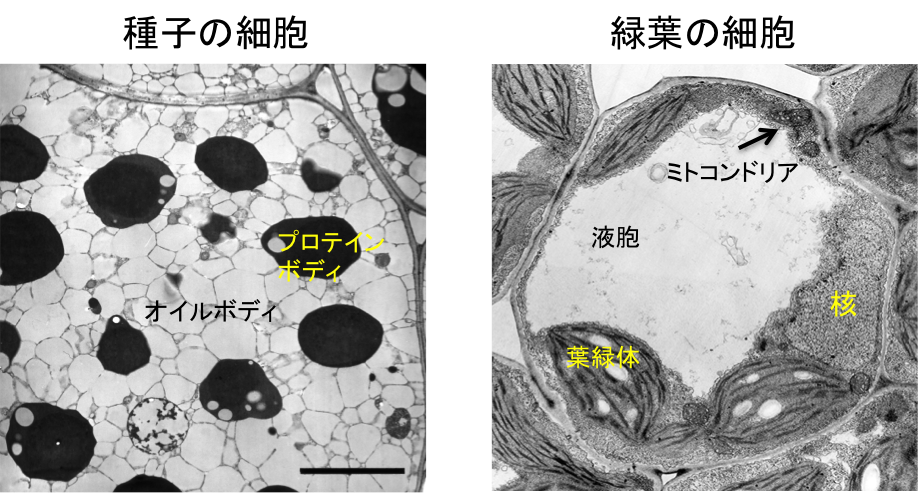
図1
2. プラスチドの分化とデンプン蓄積
プラスチドとはプロプラスチドから分化する植物に特徴的な一群の細胞内小器官です。聞いたことがないと思われるかも知れませんが、実は葉緑体もプラスチドの一種なのです。プラスチドには葉緑体の他にもアミロプラスト、エチオプラスト、ロイコプラスト、クロモプラストなど形も機能も異なるさまざまな細胞内小器官が含まれます。また、これらのプラスチドは、デンプンを蓄積する葉緑体やアミロプラストと、デンプンを蓄積しないその他のプラスチドに分類することができます。
私たちは、デンプンの蓄積という観点からプラスチド分化の制御機構を明らかにしたいと考えて、本来デンプンを蓄積しないエチオプラストにデンプンが蓄積してしまう変異体を同定しました。変異体の解析から、エチオプラストではデンプン蓄積が葉酸によって抑制されることが明らかになりました。この成果は、2018年1月17日のNHK「ガッテン」で紹介されました。
3.ペルオキシソームの機能分化
ペルオキシソームは真核生物に普遍的に存在する直径約1μmほどのオルガネラで、過酸化水素を発生型酸化酵素とそれを消去するためのカタラーゼを含むという共通の性質を持っています。高等植物の場合、環境変化などに呼応してペルオキシソームの機能が変化します。たとえば、子葉や胚乳などの細胞に存在するペルオキシソームは特にグリオキシソームと呼ばれ、脂肪酸β酸化系とグリオキシル酸回路に働きによってオイルボディに蓄えられた貯蔵脂肪をスクロースに分解する上で中心的な役割を果たしています(図2-1)。一方、光合成組織の細胞には緑葉ペルオキシソームが存在します。緑葉ペルオキシソームは、葉緑体、ミトコンドリアと強調して働き、光呼吸を行っています(図2-2)。植物が発芽・成長する過程では、同一の細胞内に存在するそれぞれのペルオキシソームが、グリオキシソームから緑葉ペルオキシソームへと機能が変化します。また、子葉が老化する過程では、緑葉ペルオキシソームからグリオキシソームへと可逆的に機能変換します。ペルオキシソームの機能変換は他の生物には見られない高等植物に特有の現象です。私たちは、植物におけるペルオキシソームの機能変換が遺伝子発現調節やタンパク質輸送、タンパク質分解、オートファジーなどの制御を受けることを明らかにしました。また、プロテオーム解析やトランスクリプトーム解析を行い、根のペルオキシソームがポリアミン分解を行うなど従来知られていた以上に多彩な機能に分化することを明らかにしています。
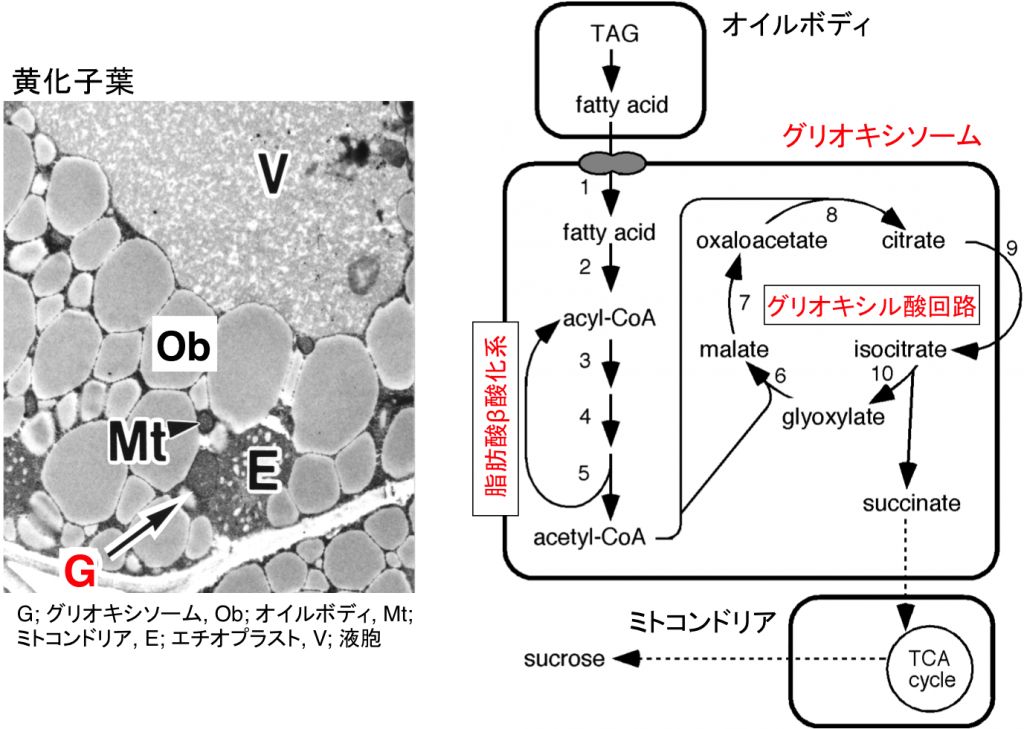
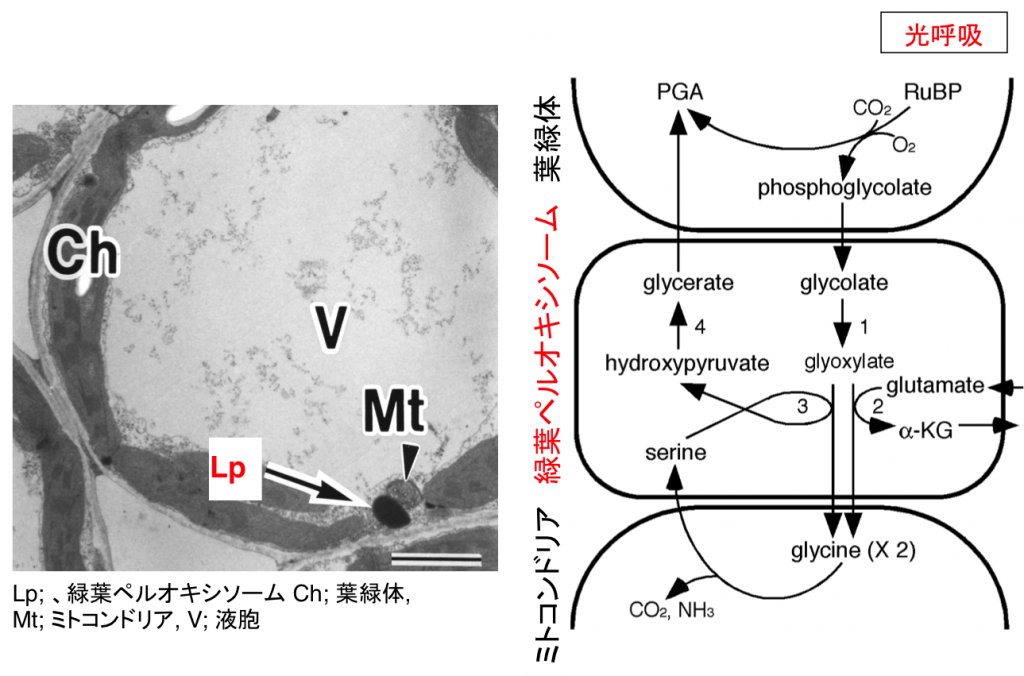
図3-1 グリオキシソームの機能 図3-2 緑葉ペルオキシソームの機能
4.ペルオキシソームのライブイメージング
ペルオキシソームタンパク質はサイトゾルのフリーポリソームで翻訳された後にペルオキシソームへと輸送されます。ペルオキシソームタンパク質はPTS1もしくはPTS2と呼ばれるペルオキシソームタンパク質輸送シグナルを含んでいます。植物の場合、PTS1はカルボキシ末端に存在する特定の組み合わせからなるトリペプチド配列、PTS2はアミノ末端の延長ペプチド内に存在する特定のアミノ酸配列であることを明らかにしています。カルボキシ末端やアミノ末端にPTS1やPTS2を付加したGFP(緑色蛍光タンパク質)融合タンパク質を発現する形質転換植物を作製することで、生きた細胞内のペルオキシソームを観察することを可能にしました(図3)。
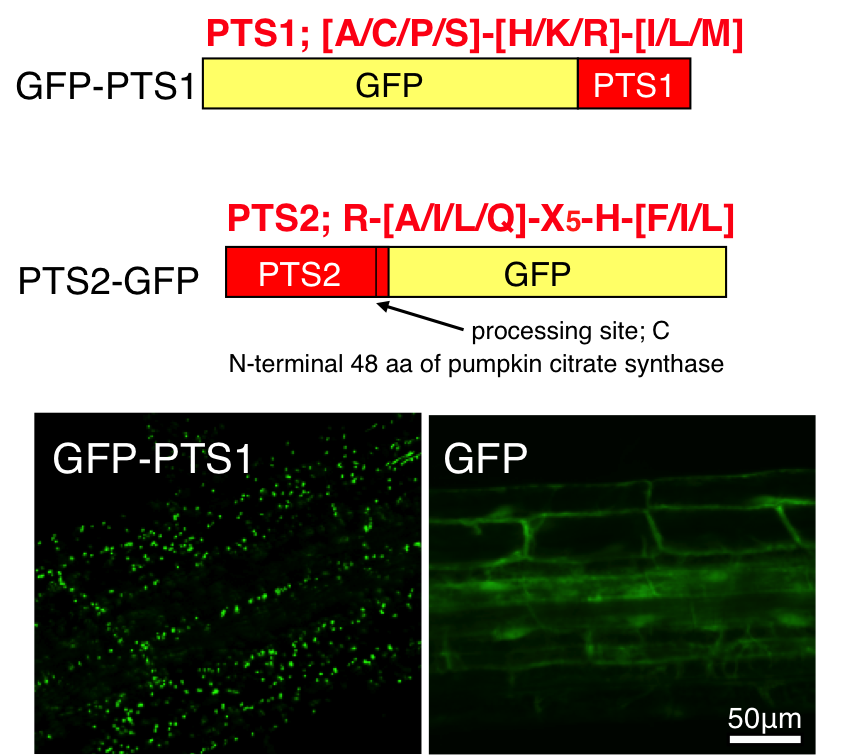
図4 ペルオキシソームのライブイメージング
5.ペルオキシソームタンパク質の輸送機構
植物のペルオキシソームタンパク質の細胞内輸送には多くのタンパク質が関与しています(図3)。ペルオキシソームタンパク質をサイトゾルで最初に識別するのはPEX5とPEX7を含むレセプターです。PEX5はPTS1と、PEX7はPTS2と結合し、PTS1-PTS2レセプター複合体を形成します。この複合体はPEX5とペルオキシソーム膜タンパク質であるPEX14が結合することで、ペルオキシソームへ運ばれます。PTS1-PTS2レセプター複合体に含まれるペルオキシソームタンパク質は、PEX13やその他のPEXタンパク質、RAbE1cなどの働きによって立体構造を保ったままペルオキシソーム膜を透過していきます。
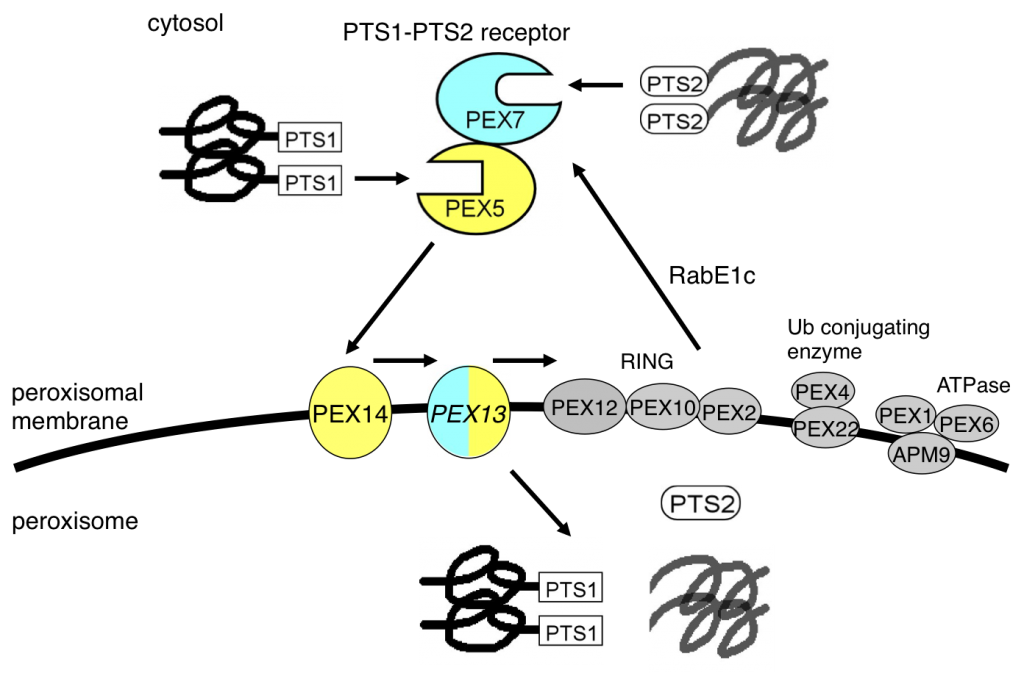
図5 ペルオキシソームタンパク質の輸送
6.グリオキシソーム機能欠損変異体
植物におけるペルオキシソームの制御機構を明らかにするために、2,4-dichlorophenoxybutyric acid(24DB)を用いてグリオキシソームの脂肪酸β酸化活性を欠損する変異体を単離しました。野生型シロイヌナズナは、24DBを添加することで成長が阻害されます。これは脂肪酸β酸化による24DBが除草剤24Dに代謝されるためです。一方、グリオキシソーム機能を欠損する変異体は24DBに耐性を示し、発芽後の成長にスクロースを要求します(図5)また、種子の発芽率が低下する変異体もあります。これら変異体を解析することでグリオキシソームが種子の発芽や発芽後の成長に果たす役割を調べています。
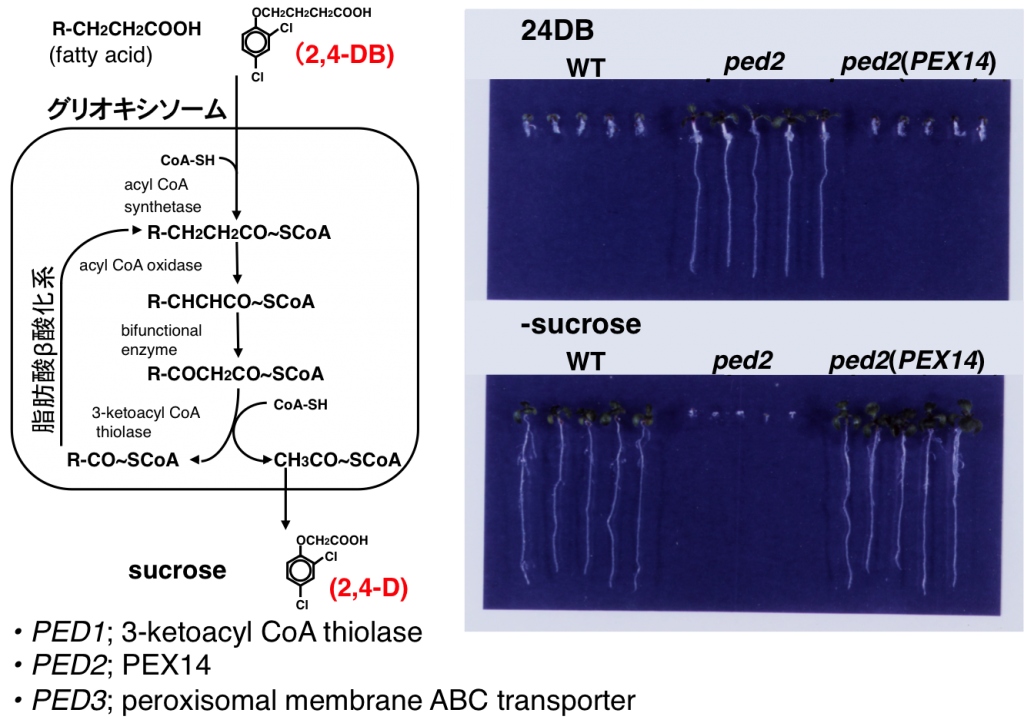
図6 グリオキシソーム機能欠損変異体の表現型
7.ペルオキシソーム形成不全変異体の単離
ペルオキシソームのライブイメージングを利用して、ペルオキシソーム形成不全変異体を作製しています(図6)。変異体の多くはペルオキシソームのタンパク質輸送機構に関わる遺伝子を欠損しており、ペルオキシソームタンパク質がサイトゾルに蓄積していました。ペルオキシソームが細長くなったり、大きくなったりする変異体も見つかりました。これら変異体の原因遺伝子はペルオキシソームの分裂や形態の維持に必須であると考えられます。
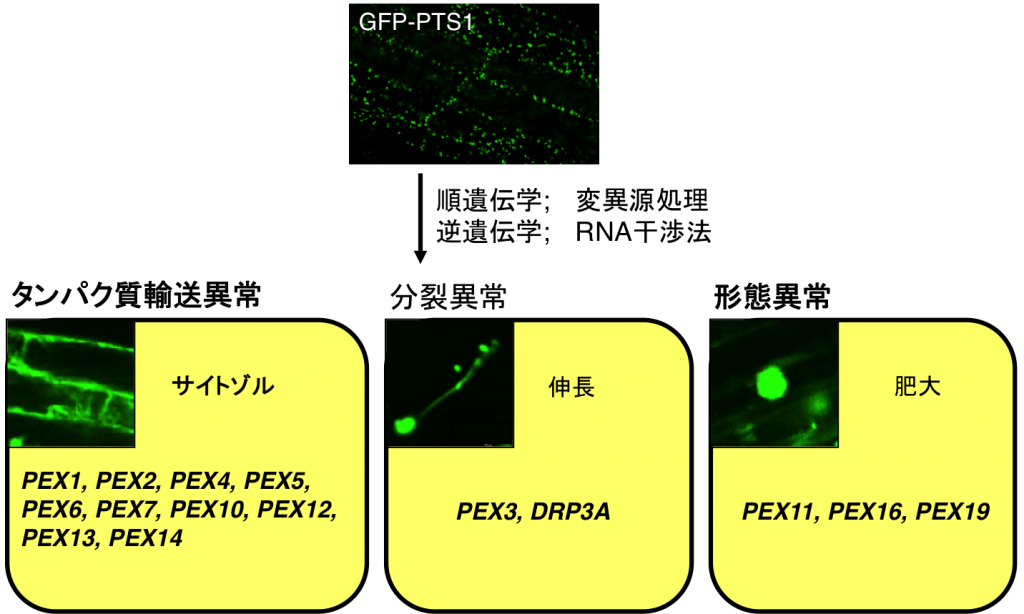
図7 遺伝子変異によって引き起こされるペルオキシソームの形成不全
8.オイルボディの形成や貯蔵脂肪の蓄積・分解のメカニズム
オイルボディは脂肪性種子の細胞に存在するオルガネラで、リン脂質の半膜とオレオシンという膜タンパク質で囲まれた特殊な構造をしています(図7)。内部には種子の貯蔵脂肪であるトリアシルグリセロールが蓄積しています。オイルボディは種子が登熟する過程で小胞体から形成されると考えられています。オイルボディの形態異常を示す変異体を単離することで、オイルボディの形成や分解機構の解明をめざしています。
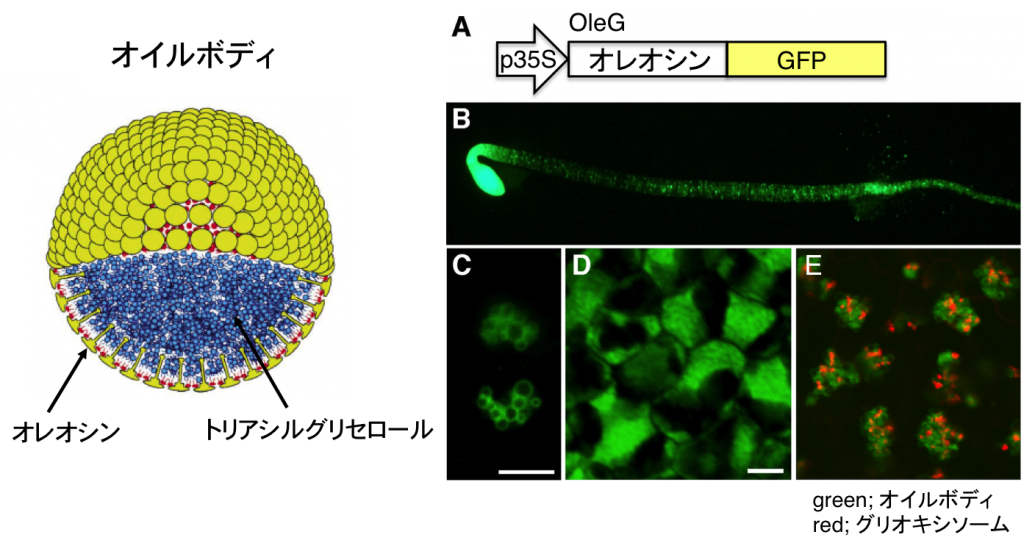
図8 GFPを用いたオイルボディのライブイメージング

